近日,土壤生态学团队在New Phytologist上发表题为“Maize growth as a function of cover crop-mediated soil microbiome”的研究论文,报道了野外大尺度土壤采样与温室盆栽实验途径相结合,进行覆盖作物(绿肥)种植调节土壤生物和非生物性质及其对玉米生长影响机制的研究。主要报道了在我国土壤初始性质变异较大的背景下、覆盖作物介导的土壤微生物调节了玉米生长的发现。研究结果有助于加强应用覆盖作物恢复和调节农田土壤生物群落和功能的认识,为将土壤生物纳入生态系统功能预测模型及进一步利用土壤生物群落促进农业生态恢复管理提供科学依据。
近年来,定向调控有益土壤生物类群、充分发挥生物调节的多功能地位已成为提升土壤健康与农业绿色发展的重要集约化生态策略(Jansson et al., 2023; Jing et al., 2022; Toju et al., 2018)。例如,微生物作为生物肥料或害虫控制的重要工具已被应用于促进作物生长,从而减少对合成化学品的依赖。然而,目前关于哪些土壤类群可提升作物生长的认知仍处于初级阶段,甚至缺乏解释大尺度下土壤微生物对作物生长相对贡献的基础认识。
当前,有关土壤微生物对作物生长影响的不确定性主要源于三个方面。首先,既有研究多聚焦局域尺度土壤微生物对作物生长的影响(Raza et al., 2024; Wang et al., 2024)。然而,目前仍不清楚在局部范围内影响作物生长的微生物群落是否也能在更大的空间尺度上影响作物生长表现。其次,现有研究多关注于少数关键微生物类群如促进作物生长的固氮菌和菌根真菌(Chakraborty et al., 2024; Lutz et al., 2023)以及威胁农业生产的常见病原菌(Liu et al., 2024)。然而,土壤生物群落极其复杂,也包括众多调节养分循环的硝化细菌、分解真菌和捕食性原生生物等,这些功能类群是否均与各种条件下的作物生长有联系,目前还不清楚。第三,众所周知,农业实践(如施肥、灌溉、耕作、轮作等)会改变土壤生物群落的组成和多样性以及养分供应和碳固存(Chaplot et al., 2023; Fan et al., 2021);特别是越来越多的研究表明,覆盖作物种植(如在主要作物休耕期间进行覆盖种植)作为一种纯粹的绿色投入品和生态调控途径,对土壤健康和植物生长都有巨大的调节潜力(Zhang et al., 2022)。目前,覆盖作物种植对土壤生物群落的影响及其如何进一步调节作物生长的机制仍不清楚。
基于此,本研究通过采集中国不同区域的27份种植典型主作物(玉米)的土壤样品,进行两阶段的温室盆栽实验(图1)。在第一阶段,种植两种典型覆盖作物,包括豆科光叶笤子和禾本科黑麦草。在第二阶段,种植主作物玉米。以此探究(1)土壤微生物群落对玉米生长的贡献是什么?(2)覆盖作物种植如何通过介导特定土壤微生物类群调节玉米生长?结果表明,土壤微生物可解释29%的玉米生长变化(图2),其中关键类群如细菌地嗜皮菌属(Geodermatophilus)、粒球菌属(Lapillicoccus)和嗜酸菌属(Acidipila)与玉米生物量呈显著正相关(图3)。此外,豆科覆盖作物种植维持了玉米生物量的稳定,这与其抑制了细菌刺戟菌属(Jatrophihabitans)和甲基小枝菌属(Methylovirgula)以及分解者真菌(Decomposer)密切相关。禾本科覆盖作物种植下玉米生物量显著降低,则主要与细菌分枝杆菌属(Mycobacterium)和分解者真菌(Decomposer)的增加有关(图4)。总的来说,本研究强调了覆盖作物种植可以通过改变特定土壤微生物类群影响后续主作物的玉米生长。这项工作加深了目前对土壤微生物群落如何促进作物生长的理解,并强调了以微生物为依据的土壤生态管理策略的潜力。
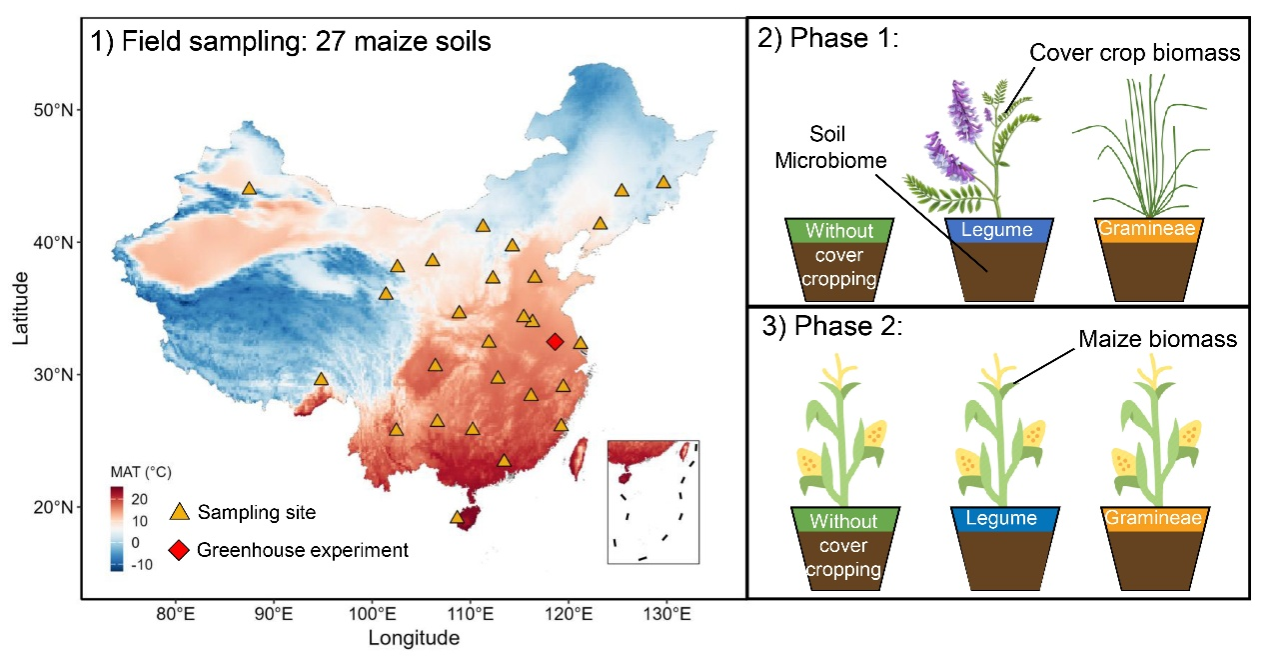
图1. 本研究的两步实验设计流程图。1)我国典型土壤的大尺度野外采样。通过采集中国不同区域的27份种植典型主作物(玉米)的土壤样品,涵盖了不同的气候和土壤条件。2)利用野外采集的土壤进行了温室盆栽实验第一阶段,并在花盆中种植两种典型覆盖作物,包括豆科光叶笤子(Vicia villosa Roth)和禾本科黑麦草(Lolium multiflorum Lam.)。在这一阶段,测量了土壤微生物、土壤酶活性、土壤碳库和氮素有效性。随后,使用相同花盆进行了温室盆栽实验第二阶段,通过种植主作物玉米,测量玉米生长的变化,包括地上生物量、地下生物量和总生物量。
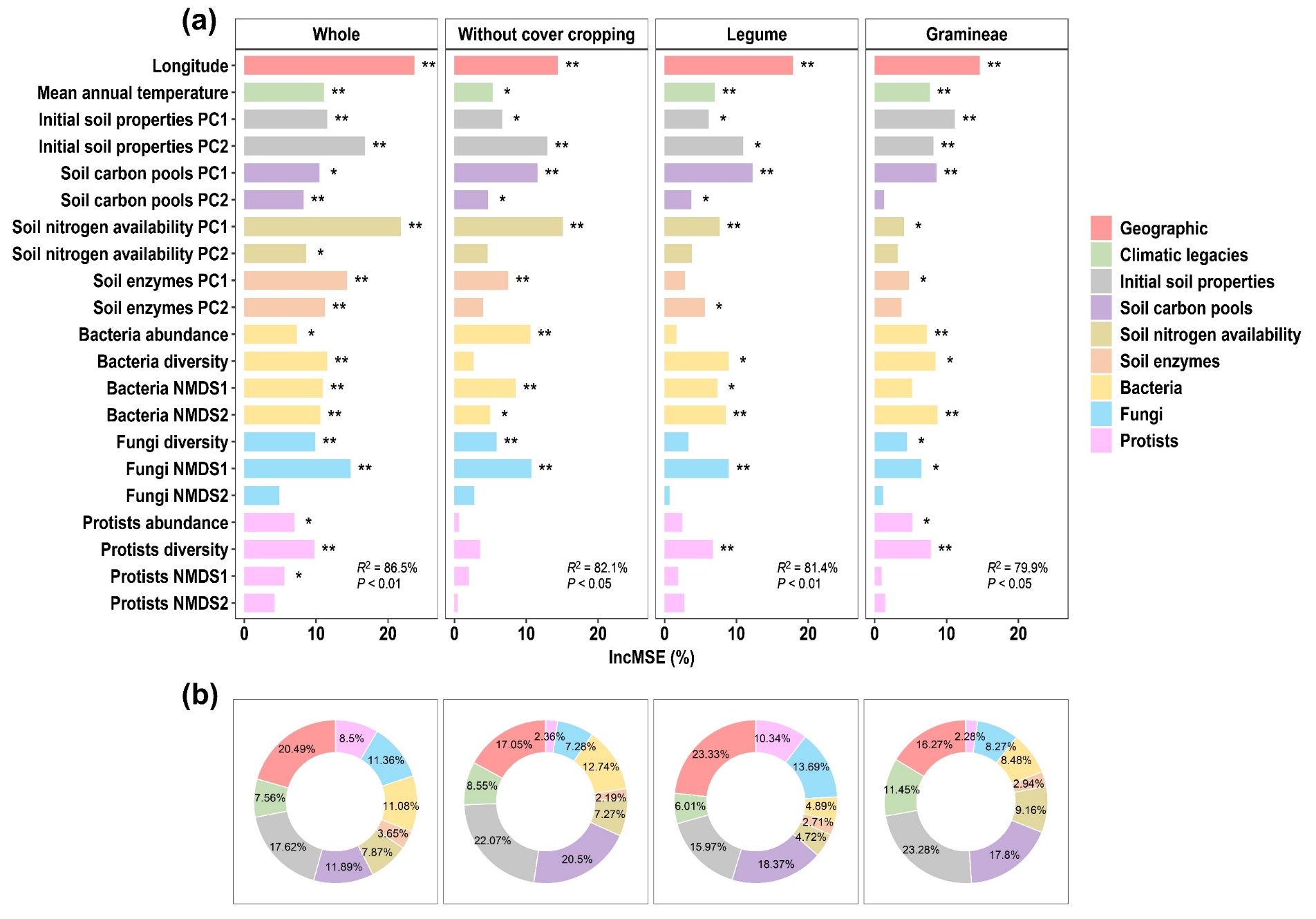
图2. 覆盖作物种植介导的生物和非生物因素对玉米生长的影响(盆栽试验)。(a) 随机森林模型(RF)显示了不同驱动因素在预测玉米生长方面的相对重要性。在随机森林模型中,均方误差百分比(IncMSE%)越高,表明特定因子的重要性越高。(b) 层次分割模型(HP)用于量化所有预测因子对玉米生长的独特贡献。选择第一主成分(PC1)和第二主成分(PC2)代表初始土壤特性、土壤有效性和土壤碳库的综合影响。微生物(细菌、真菌和原生生物)相对丰度、多样性和群落组成基于玉米相关 ASV 计算。微生物多样性以丰富度量化,而群落组成则通过基于布雷-柯蒂斯距离的非度量多维尺度(NMDS)分析的第一轴(NMDS1)和第二轴(NMDS2)进行表示。微生物相对丰度基于玉米相关 ASV 相对丰度的总和计算。
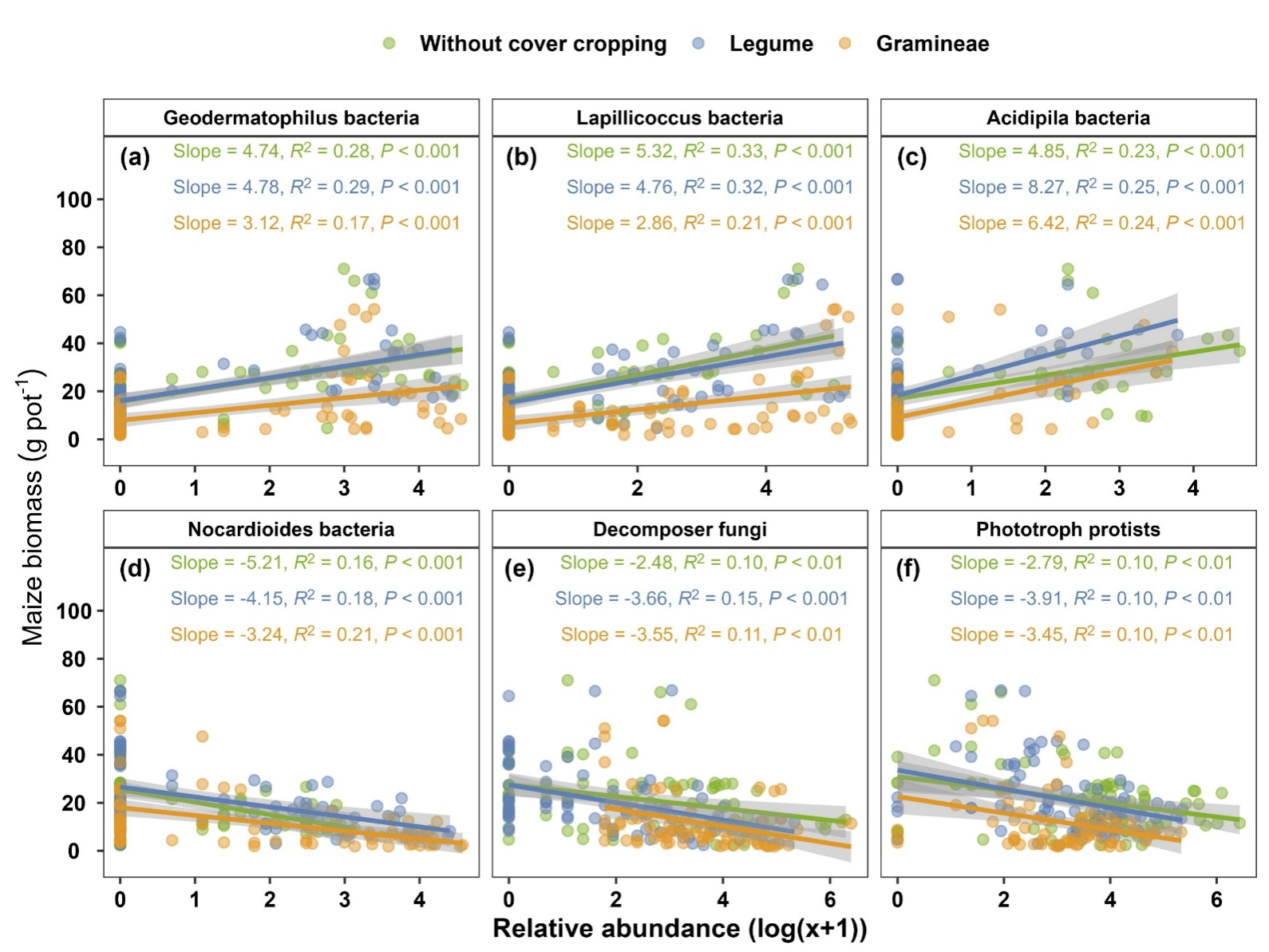
图3. 覆盖作物种植介导的土壤微生物类群与玉米生长的关系(盆栽试验)。(a-f)不同覆盖作物处理下土壤微生物类群与玉米生物量的线性回归分析。玉米生物量以总生物量表示。
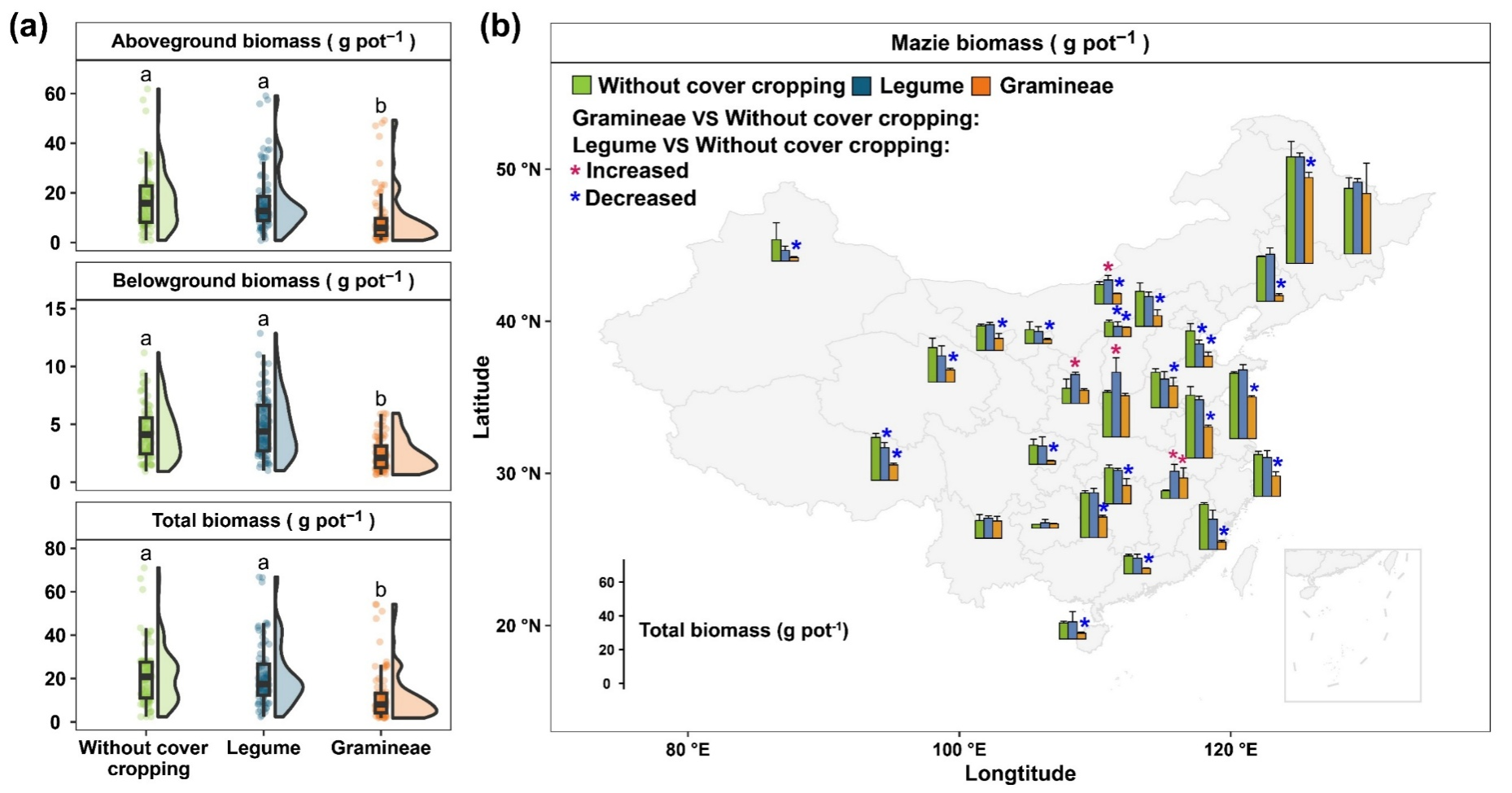
图4. 覆盖作物种植对我国不同地点采集土壤中玉米生长的影响(盆栽试验)。(a) 线性混合模型(LMMs)显示了覆盖作物种植对玉米地上部生物量、地下部生物量和总生物量的影响。在这些模型中,覆盖作物被视为固定效应,而地点和花盆则被视为随机效应。箱线图中的水平线表示中位数,箱体部分表示第25百分位数和第75百分位数,须线则表示数据的最小值和最大值。(b) 线性混合模型(LMMs)显示了覆盖作物种植对27 个地点玉米生物量的影响。在这些模型中,覆盖作物被视为固定效应,而花盆被视为随机效应。玉米生物量以总生物量表示。数值以平均值 ± SE 表示。
土壤生态实验室博士生石思博为第一作者,刘满强教授为通讯作者,西班牙塞维利亚自然资源和农业生物研究所Delgado-Baquerizo教授为共同通讯作者。本实验室陈小云教授、已毕业博士生龚鑫、程赛赛,及中山大学陶冬雪博士、瑞士苏黎世大学van der Heijden教授和美国科罗拉多州立大学Trivedi教授共同参与并指导了该项工作。本研究得到国家重点研发计划(2022YFD1500203)和甘肃省科技计划(24RCKB002)的支持。
参考文献:
Chakraborty S, Venkataraman M, Infante V, Pfleger BF, Ané JM, 2024. Scripting a new dialogue between diazotrophs and crops. Trends in Microbiology, 32(6): 577-589.
Chaplot V, Smith P, 2023. Cover crops do not increase soil organic carbon stocks as much as has been claimed: What is the way forward?. Global Change Biology, 29(22): 6163-6169.
Fan K, Delgado-Baquerizo M, Guo X, Wang D, Zhu Y, Chu H, 2021. Biodiversity of key-stone phylotypes determines crop production in a 4-decade fertilization experiment. The ISME Journal, 15(2): 550-561.
Jansson JK, McClure R, Egbert RG, 2023. Soil microbiome engineering for sustainability in a changing environment. Nature Biotechnology, 41(12): 1716-1728.
Liu H, Zhang W, He Q, Aikemu R, Xu H, Guo Z, Wang L, Li W, Wang G, Wang X, Guo W, 2024. Re-localization of a repeat-containing fungal effector by apoplastic protein Chitinase-like 1 blocks its toxicity[J]. Nature Communications, 15(1): 10122.
Lutz S, Bodenhausen N, Hess J, Valzano-Held A, Waelchli J, Deslandes-Hérold G, Schlaeppi K, Van Der Heijden MGA, 2023. Soil microbiome indicators can predict crop growth response to large-scale inoculation with arbuscular mycorrhizal fungi. Nature Microbiology, 8(12): 2277-2289.
Raza W, Jiang G, Eisenhauer N, Huang Y, Wei Z, Shen Q, Kowalchuk GA, Jousset A, 2024. Microbe-induced phenotypic variation leads to overyielding in clonal plant populations. Nature Ecology & Evolution, 8(3): 392-399.
Toju H, Peay KG, Yamamichi M, Narisawa K, Hiruma K, Naito K, Fukuda S, Ushio M, Nakaoka S, Onoda Y, Yoshida K, Schlaeppi K, Bai Y, Sugiura R, Ichihashi Y, Minamisawa K, Kiers ET, 2018. Core microbiomes for sustainable agroecosystems. Nature Plants, 4(5): 247-257.
Wang M, Ge AH, Ma X, Wang X, Xie Q, Wang L, Song X, Jiang M, Yang W, Murray JD, Wang Y, Liu H, Cao X, Wang E, 2024. Dynamic root microbiome sustains soybean productivity under unbalanced fertilization. Nature Communications, 15(1): 1668.
Zhang C, Xue W, Xue J, Zhang J, Qiu L, Chen X, Hu F, Kardol P, Liu M. 2022. Leveraging functional traits of cover crops to coordinate crop productivity and soil health. Journal of Applied Ecology 59(10): 2627-2641.
原文链接:http://doi.org/10.1111/nph.70460
编辑:石思博
校正:朱梦一,刘满强
审核:刘满强