近日,土壤生态团队在Agriculture, Ecosystems and Environment国际期刊发表题为“Nutrient limitation and saline–alkaline stress primarily drive community and function shifts in protists inhabiting saline–sodic soils”的研究论文,系统揭示了养分限制与盐碱胁迫是驱动盐碱土壤中原生生物群落结构与功能分化的主要因子,并揭示了其在盐碱土壤原生生物为核心的微食物网和生态系统功能中的核心作用。这一发现为理解全球盐渍化土壤的土壤生态学机制和盐碱地可持续利用提供了新的理论支撑。
土壤盐渍化是全球范围内最严重的土地退化类型之一,严重威胁农业生产力和生态系统稳定性。据联合国粮农组织最新数据(FAO, 2024),全球盐渍化土壤面积已超过 13.8 亿公顷,约占陆地面积的 10.7 % (Srarfi and Majar, 2024)。已有研究表明,盐渍化可深刻改变土壤细菌和真菌群落 (Hollister et al., 2010; Montanarella et al., 2015),然而,作为土壤微生物网核心的原生生物在盐碱土中的群落特征与生态功能却长期被忽视。原生生物不仅是细菌和真菌的重要捕食者,还为更高营养级(如捕杂食线虫等)提供食物来源,其在多营养级互作中的关键地位,表明将在养分循环、能量流动、病害防控和生态系统稳定性等方面发挥关键作用 (Stout, 1980; Geisen and Bonkowski, 2018; Delgado-Baquerizo et al., 2019; Gao et al., 2019)。但相较于细菌和真菌,原生生物在盐碱胁迫下的群落动态及其驱动机制研究仍极为有限 (Arndt et al., 2020; Burki et al., 2020)。
本研究针对中国松嫩平原典型盐碱化土壤,采集代表性自然苏打盐碱土壤与盐碱土开垦后的农田土壤,范围覆盖整个苏打盐碱土分布区,结合高通量测序与实时荧光定量PCR,系统分析了原生生物群落的多样性、丰度和功能分化,并耦合土壤理化性质、气候因子及其他土壤生物进行解析。研究发现,群落多样性与丰度模式相反——农田土壤中原生生物多样性更高,而自然盐碱土中绝对丰度更高;功能群显著分化——消费者类群在盐碱土中占优势,而光合型原生生物主导农田土壤(图1, 2);主要环境驱动因子明确——总氮(TN)和总磷(TP)等养分指标,以及交换性钠百分比(ESP)、pH 等盐碱胁迫指标,是决定原生生物多样性和群落结构的关键因子,而气候变量作用较弱;跨营养级互作影响也达到显著水平——原生生物群落与细菌丰度、线虫(食细菌和食真菌)高度相关,凸显其在土壤微食物网中的核心作用(图3, 4),这也验证了先前的报道 (Geisen, 2016; Gao et al., 2019)。

图 1. 自然苏打盐碱土壤与改良农田土壤中原生生物群落的比较
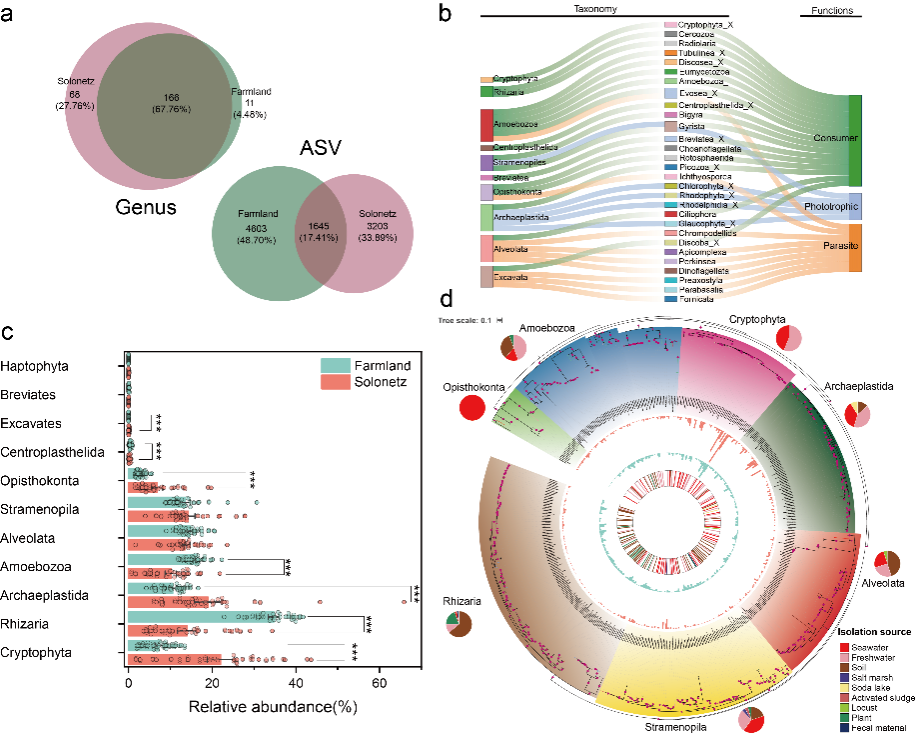
图2. 自然苏打盐碱土壤与改良农田土壤中原生生物群落结构、多样性及功能分化的比较分析
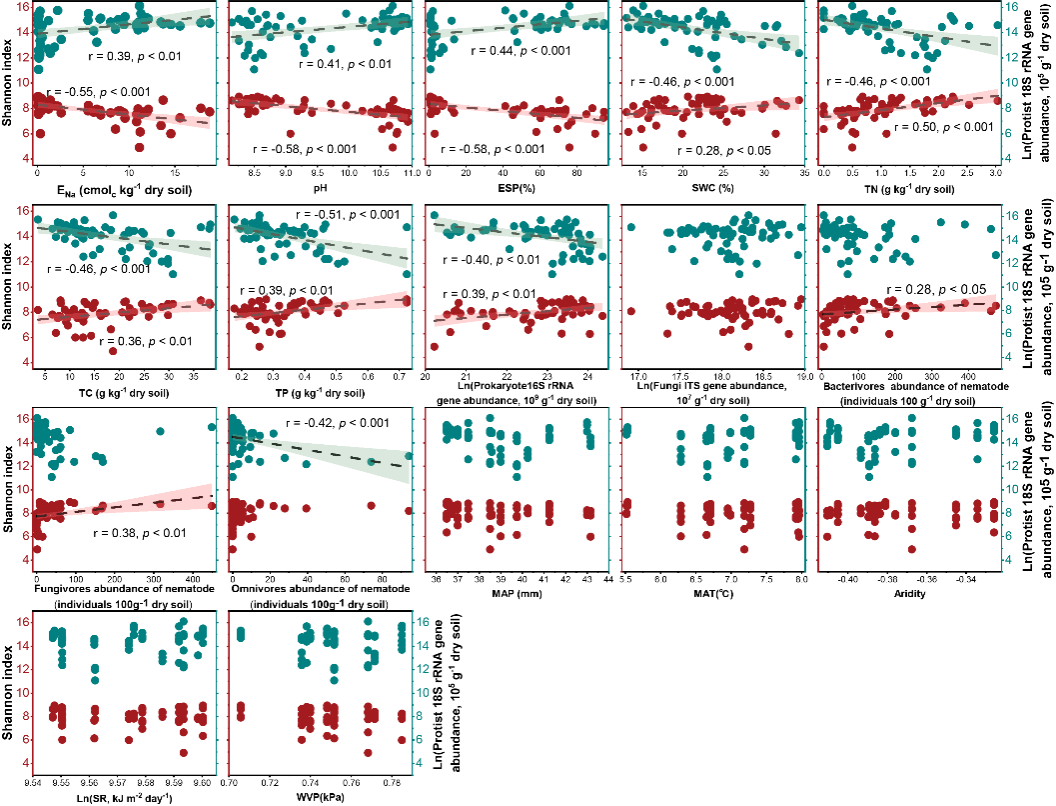
图3. 原生生物多样性和丰度与主要土壤理化、生物(微生物与土壤动物)及气候因子之间的相关关系
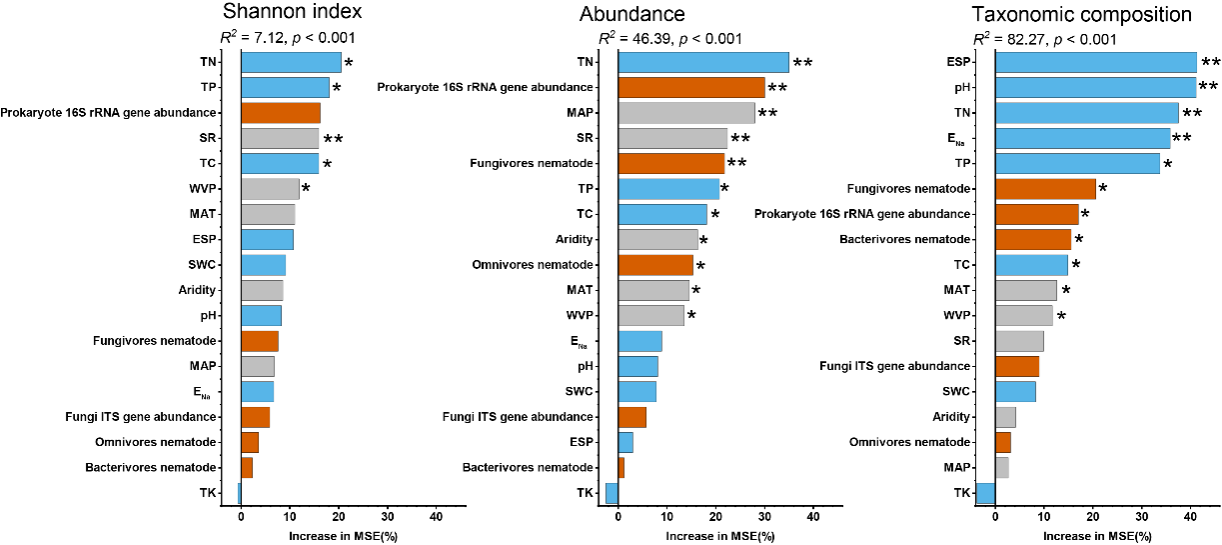
图4. 盐碱胁迫下,土壤理化、生物及气候因子对塑造土壤原生生物多样性、丰度及群落组成的相对重要性
研究结果表明,盐碱胁迫与养分限制共同塑造了原生生物群落和生态功能格局,而长期农田开垦能够有效缓解盐碱胁迫,提升原生生物多样性,推动土壤功能向更高稳定性和多功能性转变。这一发现不仅拓展了对盐碱土壤微食物网的认识,也为盐碱土壤改良与管理提供了重要理论依据。
土壤生态实验室青年研究员孙祥鑫博士为第一作者,刘满强教授为通讯作者,本实验室陈小云教授参与并指导了该项工作。研究得到国家重点研发计划(2022YFD1500203)、国家自然科学基金(42407415)、江苏省基础研究专项(BK20241555)、中国博士后科学基金(2022M721645)和江苏省卓越博士后计划(2023ZB244)的联合资助。
主要参考文献
Arndt, H., Ritter, B., Rybarski, A., Schiwitza, S., Dunai, T., Nitsche, F., 2020. Mirroring the effect of geological evolution: Protist divergence in the Atacama Desert. Global and Planetary Change 190.
Burki, F., Roger, A.J., Brown, M.W., Simpson, A.G.B., 2020. The New Tree of Eukaryotes. Trends in Ecology & Evolution 35, 43-55.
Delgado-Baquerizo, M., Bardgett, R.D., Vitousek, P.M., Maestre, F.T., Williams, M.A., Eldridge, D.J., Lambers, H., Neuhauser, S., Gallardo, A., García-Velázquez, L., Sala, O.E., Abades, S.R., Alfaro, F.D., Berhe, A.A., Bowker, M.A., Currier, C.M., Cutler, N.A., Hart, S.C., Hayes, P.E., Hseu, Z.Y., Kirchmair, M., Peña-Ramírez, V.M., Pérez, C.A., Reed, S.C., Santos, F., Siebe, C., Sullivan, B.W., Weber-Grullon, L., Fierer, N., 2019. Changes in belowground biodiversity during ecosystem development. Proceedings of the National Academy of Sciences of the United States of America 116, 6891-6896.
Gao, Z.L., Karlsson, I., Geisen, S., Kowalchuk, G., Jousset, A., 2019. Protists: Puppet Masters of the Rhizosphere Microbiome. Trends in Plant Science 24, 165-176.
Geisen, S., 2016. The bacterial-fungal energy channel concept challenged by enormous functional versatility of soil protists. Soil Biology & Biochemistry 102, 22-25.
Geisen, S., Bonkowski, M., 2018. Methodological advances to study the diversity of soil protists and their functioning in soil food webs. Applied Soil Ecology 123, 328-333.
Hollister, E., B., Engledow, A., S., Hammett, A., J., Provin, 2010. Shifts in microbial community structure along an ecological gradient of hypersaline soils and sediments. The ISME Journal 4, 829.
Montanarella, L., Chude, V., Yagi, K., Krasilnikov, P., Panah, S.K.A., Maria de Lourdes Mendonça-Santos, M.L., Pennock, D., McKenzie, N., 2015. Status of the World's Soil Resources (SWSR) - Main Report.
Srarfi, F., Majar, A., 2024. Global Status of Salt-affected soils, pp. 83-107.
Stout, J.D., 1980. The Role of Protozoa in Nutrient Cycling and Energy Flow, In: Alexander, M. (Ed.), Advances in Microbial Ecology. Springer US, Boston, MA, pp. 1-50.
原文链接:https://doi.org/10.1016/j.agee.2025.110009
编辑:孙祥鑫
校正:朱梦一,刘满强
审核:刘满强